

Abstract
Adults diagnosed with acute myeloid leukemia have a poor prognosis, with only 30% surviving at five years. Despite advances in our understanding of the molecular underpinnings of AML, the chemotherapy backbone used to treat most patients (combination cytarabine and anthracycline) remains unchanged since 1973.
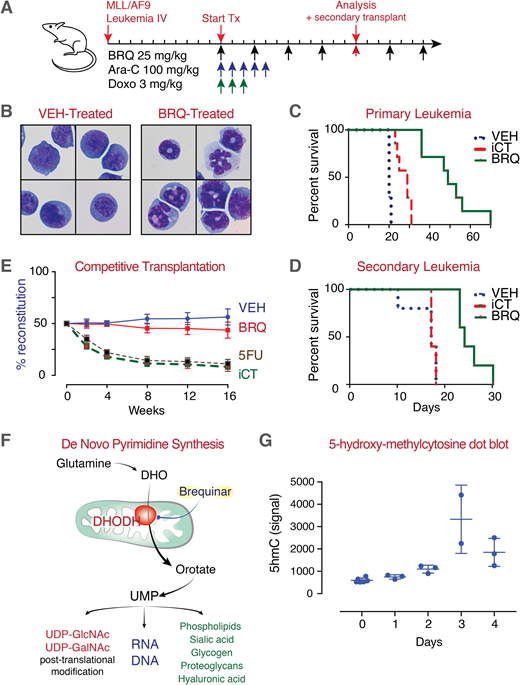
With the goal of identifying a new differentiation therapy, we previously performed a small molecule phenotypic screen to find compounds that triggered myeloid maturation. Unexpectedly, our most active hits were inhibitors of the enzyme dihydroorotate dehydrogenase (DHODH). The DHODH-inhibitor brequinar demonstrated dramatic activity in vitro and in vivo (Figure 1A), leading to differentiation (Figure 1B) and prolonged survival across multiple AML models when compared to standard induction chemotherapy (iCT) (Figure 1C). Brequinar treatment also depleted leukemia initiating cells in vivo as measured by secondary transplant analysis (Figure 1D).
Cancer therapy requires a 'therapeutic window' such that the treatment must target and kill malignant cells before killing normal cells and causing unacceptable side effects. An essential and ubiquitously expressed enzyme, DHODH is not typical among most cancer drug targets and is not known to be mutated or overexpressed in malignant cells. Yet empirically, a DHODH-inhibitor therapeutic window exists.
The competitive transplantation of hematopoietic stem cells following 36-days of brequinar treatment (at the same dose that is anti-leukemic and pro-differentiation) does not lead to a competitive disadvantage or deleterious differentiation of normal stem cells (Figure 1E). The basis of this therapeutic window in the case of DHODH inhibitors is not known.
Here we address the question: how are malignant cells metabolically programmed such that they are preferentially sensitive to pyrimidine starvation following inhibition of DHODH? Uridine is involved in myriad cellular processes beyond DNA and RNA synthesis, implicating many metabolic pathways (Figure 1F). We are taking an approach of (1) measuring metabolite levels and (2) metabolic pathway flux, as well as (3) using small molecule and genetic perturbations (shRNA) to unravel the differences in pyrimidine metabolism between normal and malignant cells.
The mechanism through which treatment with brequinar and inhibition of pyrimidine synthesis results in myeloid differentiation is also not known. Our preliminary data suggest that brequinar treatment not only leads to a rapid depletion of pyrimidines and downstream metabolites, but also to changes in TCA cycle metabolites including succinate and fumarate. Given the importance of these metabolites in the regulation of the TET-family of proteins, we measured changes in 5-hydroxy-methylcytosine as a surrogate for TET-enzyme activity following treatment with brequinar (Figure 1G). An increase in global 5hmC, suggesting an increase in TET-enzyme activity, paralleled the cellular differentiation over the first 72-hours. This is an intriguing association given the importance of TET2 in myeloid differentiation and the observation that inhibition of TET2 contributes to myeloid differentiation arrest.
As of summer 2018, two small molecule DHODH inhibitors are in early stage human clinical trials with two more inhibitors poised for 2019. A careful understanding of these metabolic pathways is important to inform the clinical experience of this class of anti-metabolite therapeutics. Understanding the basis of this therapeutic window will be critical to the safe and effective use of DHODH inhibitors in patients with advanced hematologic malignancies.
Vander Heiden:Agios: Membership on an entity's Board of Directors or advisory committees; Aeglea: Membership on an entity's Board of Directors or advisory committees. Sykes:Clear Creek Bio: Equity Ownership, Other: Co-founder.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal